
|
Woody detritus is an important, but overly neglected part of forested ecosystems. To date its role in carbon dynamics has received little attention at either the local or global scale, but recent studies indicate much of the C efflux associated with land-use change results from woody detritus combustion and decomposition. Unfortunately the decomposition rates used in these analyses do not have a firm empirical or conceptual basis. It is quite clear that major uncertainties about the amounts of carbon released by forest disturbances and the total carbon stores of forest ecosystems will not be resolved until woody detritus decomposition rates are either measured directly or predicted using a mechanistic basis.
To provide empirical data we are now measuring rates at 14 sites that cover the entire range of climatic conditions in which forests grow. These include nine sites dominated by conifers (Andrews-OR, Ashland RNA-OR, Cascade Head-OR, Fraser-CO, Medicine Bow-WY, Mount Rainier-WA, Pringle Falls-OR, Sequoia-CA, Wind River-WA), one conifer-hardwood mixed site (St. Petersburg, Russia), a temperate hardwood dominated site (Chapel Hill, NC) and three Mexican tropical hardwood sites (Los Tuxtlas, Puerto Morelos and Nobeck). Tree genera available at these sites include:
|
|
This gives us over 60 species to examine.
We are examining the following:
1) The interaction between species decay resistance and size. We hypothesize that the species with the most decay resistant heartwood will have the greatest decrease in decomposition rate with increases in diameter.
2) The interaction of macroclimate and species. We hypothesize that as the macroclimate becomes more favorable to decomposition, the species with the least decay resistance have the greatest response.
3) The interaction between macroclimate, size, and position of the pieces. Based on our prior observations we hypothesize that under the driest macroclimate the smallest, suspended material decomposes slowest, but under the wettest macroclimatic conditions the largest downed material decomposes the slowest.

For each site a range of piece sizes from 1 cm diameter to as large as is available is being used to determine decomposition rates. We use size classes on a logarithmic scale (i.e., 1, 2, 4, 8, 16, 32 cm). In the species related experiments (1 and 2 above), the least- and most-decay resistant species available at each site were placed on the surface of the forest floor under closed canopies. In the macroclimate, size and position interaction experiment we placed pieces so that some are suspended off the forest floor, some are in contact with it, and some are buried. Sampling intervals depends on the site; in tropical sites samples will be collected at 6 month to 1-year periods and at the other sites 1-2 year intervals are employed.
To set up the studies of wood <15 cm in diameter we cut fresh branch material into lengths approximately 10 times the diameter of the size-class. After determining the fresh weight, subsamples were used to determine the initial moisture content, the cut ends of each piece will be sealed with neoprene or latex paint, and numbered aluminum tags attached with UV resistant cable ties. Pieces that were buried are wrapped in 1 mm nylon mesh to aid in recovery. Suspended pieces are held off the ground by means of a ladder-like arrangement of nylon strings hung from tree boles.
For wood >15 cm in diameter we sampled the least and most decay resistant species at a minimum. Our primary approach in these extensive studies is to use the chronosequence method to estimate the mean rate of decomposition. Dates of log formation are taken from long?term permanent plot records, tree scars, and other dating methods. We estimate fragmentation losses by comparing current volume to the original based on DBH at time of death. Sampled logs are "dissected" using a chainsaw. The depth of staining, advanced decay, and extent of termite galleries are noted, and 3 to 4 cross-sections are removed to provide samples for density, volume of insect galleries, C chemistry and N content. Density is determined from the volume and weight of the entire cross-section in the field and removing subsamples to determine moisture content. While the chronosequence method is not as precise as a time series, it provides good preliminary estimates of decomposition rates. We are increasing the precision of these estimates by resampling the logs in these chronosequences to produce a hybrid between time series and chronosequence methods (Harmon et al. 2000). Depending on the site, a 3 to12 year period between sampling times of logs has occurred, long enough for there to be a detectable change in mass.
An example of this study comes from North Carolina. In 1996, a long-term experiment to determine the decomposition rate of wood was established in forests of the North Carolina Botanical Garden, just after Hurricane Fran had passed through. Two species, Carya tomentosa and Quercus alba, were examined to see trend in mass loss over time. Each year various sized pieces of wood ranging from 1 to 8 cm diameter have been removed, oven dried and then weighed to determine the mass remaining (Figure 1). Mass loss has been steady throughout the experiment for these two species; however, the first year appeared to be slower than the sequent ones. This lag in decomposition was probably caused by the time it took decomposer fungi to colonize the wood. We anticipate rates of decomposition of species with decay-resistant heartwood to eventually slow as the experiment progresses.
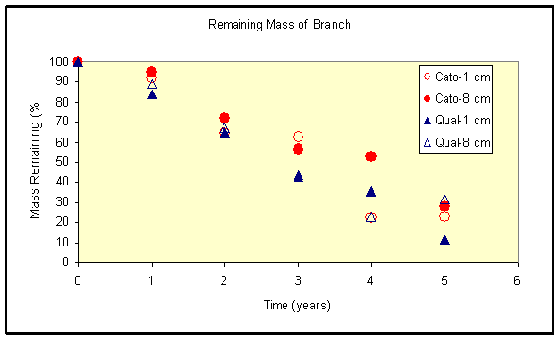 |
Figure 1. Changes in mass remaining as wood decomposition proceeds at North Carolina Botanical Garden. |
Not all wood decomposes at the same rate, and differences associated with the chemical properties of various species can have a major influence. A total of eleven species were examined, with Acer rubrum, Carya tomentosa, Fagus grandifolia, Lirodendron tulipifera, Liquidambar styraciflua, Quercus alba, and Quercus rubra having at little as 15% of their mass remaining after 4 years (Figure 2). In contrast, Juniperis virginiana and Sasafrass albidum had over 40% of their mass remaining after the same period. The latter two species have very decay-resistant heartwood with high concentrations of fungitoxic compounds. Pinus echinata and P. taeda, both with resins in their heartwoods are intermediate between these two groups of species.
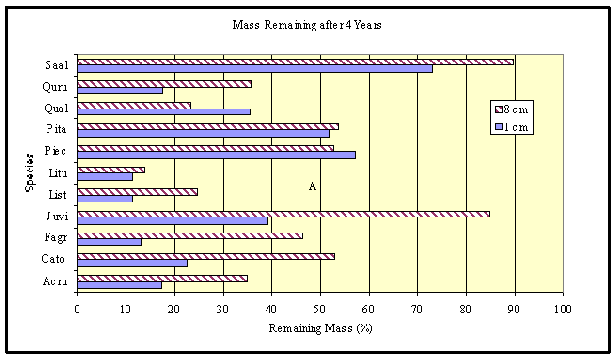 |
Figure 2. Differences in species observed in North Carolina Botanical Gardens wood decomposition experiment. |
Intuitively it makes sense that larger diameter wood would take longer to decompose than small pieces. There is some evidence for this effect, although it is not as profound as one would expect for some species (Figure 3). For example, for species with little decay-resistant heartwood, such as C. tomentosa, L. tulipifera, and Q. alba, there is a slight increase in mass remaining at 4 years as piece diameter increases. In contrast, for species with decay-resistant heartwood, such as J. virginiana, the mass remaining increases dramatically as diameter increases.
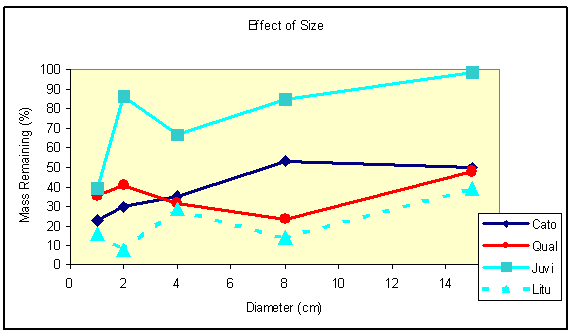 |
Figure 3. Change in mass remaining after 4 years in response to piece diameter. |
Hurricane Fran added dead wood to many microclimates ranging from suspended in the air to placed on the soil surface to in the soil. In the North Carolina climate, suspended wood dries faster than wood on the soil surface or wood in the soil. This is reflected in the mass remaining after 4 years, with suspended wood having the slowest decomposition (Figure 4). Wood decomposing on the soil surface or within the soil itself were quite similar, although we expect to see local site differences as one Mason Farm site has extremely moist samples that might slow decomposition because of waterlogging.
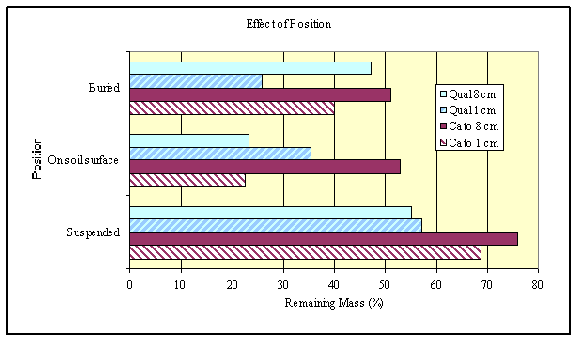 |
Figure 4. Effect of position of decomposing wood relative to the soil surface on mass remaining after 4 years of decomposition. |
We have also been using these data and those from the literature to estimate the store of dead wood at the global-level. In recent years, methods to study the size and dynamics of these detritus pools have been developed and applied to various ecosystems. Despite an increase in plot-level efforts, no reliable inventory-based estimates exist at the regional, national, or global scales. We used a series of complementary methods to estimate potential steady-state and actual stores at regional to global scales. These include: (1) correction and conversion factors for incomplete inventories, (2) dead:live wood expansion factors, (3) predictions of steady-state stores from input:decomposition rate ratios, and (4) adjustments to include disturbance regimes (Harmon et al. 2001). These methods indicate that global stores of carbon in woody detritus could be 114 to 157 Pg in the absence of disturbance, 131 to 148 Pg with natural disturbance, and 114 to 129 Pg after pre-1950's deforestation is considered. These estimates of woody detritus stores are quite crude, but they strongly indicate several points. Considerable woody detritus is present at the global level, with the bulk of it stored in evergreen forests. Globally these estimates exceed those for surface litter (50-60 Pg C) and are roughly half of that estimated for peat. Although smaller than the commonly accepted store for soils ("1500 Pg C), woody detritus is highly sensitive to natural disturbance and management by humans. This analysis indicates better inventories of dead woody detritus and measurement of its dynamics would considerably improve the overall understanding the role of carbon stores in the terrestrial biota.
We will be continuing to maintain these experiments, seeking additional funding for this effort. In addition we would like to establish experiments in environments drier than those capable of supporting closed forests (e.g., woodlands).
Harmon, M. E.; Krankina, O. N.; Yatskov, M.; Matthews, E. 2001. Predicting broad-scale carbon stores of woody detritus from plot-level data. In: Lal, R.; Kimble, J. M.; Follett, R. F.; Stewart, B. A., eds. Assessment methods for soil carbon. Boca Raton, FL: Lewis Publishers/CRC Press LLC: 533-552. (Pub No: 2806)
Harmon, M. E., O. N. Krankina, and J. Sexton. 2000. Decomposition vectors: A new approach to estimating woody detritus decomposition dynamics. Canadian Journal of Forest Research. 30: 74-84. (Pub No: 2598)
Original document posted February 18, 2002